エーギロカシス
エーギロカシス(Aegirocassis[2])は、約4億8,000万年前のオルドビス紀に生息したラディオドンタ類の節足動物の一属。2メートルの巨体に長大な甲皮と櫛状の前部付属肢をもつ[2]、モロッコで見つかった Aegirocassis benmoulai [4]という1種のみによって知られている。カンブリア紀以外の地質時代に生息した数少ないラディオドンタ類の一つである[5]。
| エーギロカシス | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 エーギロカシスの復元図
| |||||||||||||||||||||||||||
| 地質時代 | |||||||||||||||||||||||||||
| 古生代オルドビス紀前期 (約4億8,800万 - 4億7,200万年前[1]) | |||||||||||||||||||||||||||
| 分類 | |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| 学名 | |||||||||||||||||||||||||||
| Aegirocassis Van Roy, Daley, & Briggs, 2015 [2] | |||||||||||||||||||||||||||
| タイプ種 | |||||||||||||||||||||||||||
| Aegirocassis benmoulai [4] Van Roy, Daley, & Briggs, 2015 [2] |
名称 編集
学名「Aegirocassis」は、北欧神話の海神エーギル(Ægir)とラテン語の「兜」(cassis)の合成語であり、巨大な姿と発達した甲皮に因んで名付けられた[2]。模式種(タイプ種)の種小名「benmoulai」(原記載では「benmoulae」と表記された[2])は化石収集家 Mohamed Ben Moula への献名[2]。
化石 編集

エーギロカシスの化石標本は、モロッコのオルドビス紀前期(トレマドキアン期 - フロイアン期、約4億8,800万 - 4億7,200万年前[1])に当たる堆積累層 Fezouata Formation(Fezouata biota)のみから発見されている[2]。Van Roy et al. 2015 により正式に命名と記載をなされたが、それ以前の Van Roy & Briggs 2011 に記載されたオルドビス紀の巨大ラディオドンタ類の胴部化石標本 YPM 226437[1]も本属のものである[2]。また、通常のラディオドンタ類のような扁平につぶされたものとは異なり、エーギロカシスの化石は、いずれも良好な立体状態が保存されている[2]。ただし眼と口(歯)の部分は発見できず、形態は不明[2]。化石標本は、アメリカ合衆国コネチカット州ニューヘイブンのイェール・ピーボディ自然史博物館(Yale Peabody Museum of Natural History)に所蔵されている[2]。
形態 編集

-
 エーギロカシスの全身復元図。眼と尾部は推測的である。
エーギロカシスの全身復元図。眼と尾部は推測的である。
エーギロカシスは知られている中では最大のラディオドンタ類である[6]。既知最大の全身化石だけでも1.3m以上で、その比率(体長は背側の甲皮長の2倍)にあわせて単離した既知最大(1m)の背側の甲皮から換算すると、最大の体長は2mにも及ぶ[6]。これにより、本属は一部の大型ウミサソリやアースロプレウラと並んで最大級の節足動物ともなる[2]。
頭部 編集


-
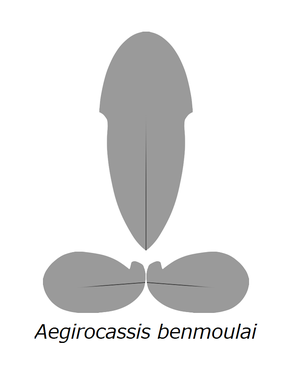 エーギロカシスの甲皮
エーギロカシスの甲皮 -
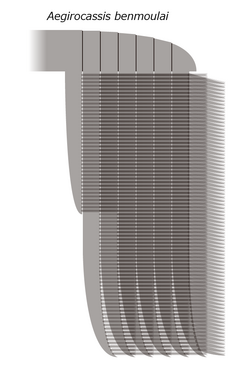 エーギロカシスの前部付属肢
エーギロカシスの前部付属肢 -
 エーギロカシス(A)とスードアングスティドントゥス(B)の前部付属肢比較図
エーギロカシス(A)とスードアングスティドントゥス(B)の前部付属肢比較図



他の多くのフルディア科のラディオドンタ類に似て、エーギロカシスの頭部には3枚の巨大な甲皮がある。雫型の背側の甲皮(H-element)は体長の半分を占めるほど長く、背側の正中線は前端ほど明瞭に隆起する[2]。その両縁には下向きの三角形の突起があり、下にある左右の甲皮の先端に連結する部分であったと考えられる[2]。左右の甲皮(P-element)は楕円形で、それぞれの前端は前述の三角形の突起に噛み合うような溝がある。表面も背側の甲皮のように、それぞれの正中線辺りにも前端ほど明瞭な隆起線がある[2]。
前部付属肢(frontal appendage)の肢節部分は約12cmで7節の肢節(柄部1節と残り6節)に分かれ、体に対して貧弱である。しかし第2- 7肢節の腹側には、肢節の長さを超えるほどの細長い6本のブレード状の内突起(endite)がある[2][3]。その先端は内側に湾曲するが、シンダーハンネスに似て、湾曲は控え目で他のフルディア科の種類より直線的である[7]。それぞれの内突起の内側には、長い剛毛様の分岐(auxiliary spines)が80本ほど並んでおり、基部は丸い関節に分かれている[2]。第2-6肢節の場合、それぞれの分岐は更に2列の繊毛が全長でV字状に並んでいる[2]。第7肢節の場合、分岐は比較的丈夫で繊毛は背面1列のみである[3]。第1肢節は第2-6肢節と似た内突起をもつが、明らかに短い[2]。また、前部付属肢の化石のうち内突起は肢節よりも保存状態が良好のため、内突起が肢節よりも頑強であったと考えられる[2]。
似たような前部付属肢は同じエーギロカシス亜科のスードアングスティドントゥスにも見られるが、そちらは内突起の分岐が対になることと繊毛を持たないことでエーギロカシスから区別できる[3]。
原記載で全身復元図を作成されたが、未発見の眼と口(歯)の形態は不明である[2]。しかし他の柔軟な部位も比較的に良好な保存状態をもつため、眼と歯は保存されにくいほど貧弱であったと推測されている[2]。また、濾過摂食者(後述)であったため、発達した視力や頑強な歯の必要性は低かったと考えられる[2]。
胴部 編集
胴部は背腹に扁平で尾部を除いて10節からなり、前後の幅の変化は控え目である[8]。第3胴節で最も幅広く、後方ほど幅狭くなる[8]。各胴節は2対の鰭(ひれ)を有し、箱型の胴節の四隅に配置される[2]。背側の鰭(dorsal flap)はサメの背びれに似た形だが、腹側の鰭(ventral flap)は丸みを帯びた三角形で、背側の鰭より少し発達していた[2]。背腹の鰭の付け根は隣接せず、側面の体表によって上下に分かれている[2]。他のラディオドンタ類に比べれば、エーギロカシスの鰭は胴部に対して小さく、背側の鰭だけでなく、腹側の鰭も他のラディオドンタ類に見られる重なり構造はない[2]。また、腹側の鰭は全長を走る脈(tranverse rods、strengthening rays)が密生しており[8]、それぞれ無数の幅狭い管状構造に細分され、おそらく筋組織をもっていたと思われる[2]。背側の鰭もそれに似たような細い脈をもつ[2]。
他のラディオドンタ類と同様、各胴節の背面には鰓と思われる一連の櫛状の構造体(setal blades)が配置される[1][2]。この構造体は左右が背側の鰭の付け根に連結し、残りの部位は胴部の背側から分離していたと思われる[2]。それを構成する細長い葉状の附属体(lanceolate blade)は、前端近くから管状の内部構造に貫通され、左右両面には皺が密生する[2]。ペイトイアと同様、この構造体は背面全体を覆い、左右に分かれていない[2]。
知られる化石標本では尾部の保存状態が悪く、Van Roy et al. 2015 ではペイトイアに似た単調な鈍い突起と解釈された[2]が、附属体(尾扇・尾毛など)の有無は不明である[6][8]。
生態 編集

エーギロカシスは穏やかに遊泳しながら、水中の動物プランクトンを摂る濾過摂食者であったと考えられる(後述)[2]。また、断片化石の密集した保存状態、サイズの多様性や三葉虫などの底生性動物も化石に含まれていたことにより、これらの化石は脱皮殻であり、エーギロカシスは海底で脱皮を行っていたことも示唆される[2]。
移動と呼吸 編集
鰭に見当たる脈の高い密度とそれを構成した大量の管状構造は、エーギロカシスはこれらの鰭の動きと形を幅広く操作できた機敏な遊泳性動物であったことを示唆する[2]。大きな甲皮・小さな鰭・背腹に扁平の体・繊毛が生えた長い内突起をもつ前部付属肢などの形質は、一定の速度での長距離の遊泳に最適化されており、急加速や高速遊泳には向いていなかったと思われる[2]。また、エーギロカシスは背腹2セットの鰭をもっているが、上下の鰭を連動すると渦が重ねて推進力が妨げられるため、腹側の鰭のみ遊泳に用いられたと思われる[2]。背側の鰭は遊泳に用いられず、代わりに遊泳中にステアリングやバランスを安定させる機能をもっていたと考えられる[2][8]。
他のラディオドンタと同様、背側の鰭の付け根に連結しながら胴節の背面を覆いかぶさった setal blades は、ガス交換およびイオン交換(両方またはいずれか一方)に用いられた呼吸器官(鰓)であったと考えられる[2]。それを構成する葉状附属体の柔軟性・細長い形・表面積を増やした両面の皺などの形質は、この機能に裏付ける証拠とされている[2]。
濾過摂食の仕組み 編集
前部付属肢の繊毛の間隔から推測すると、エーギロカシスは同じ懸濁物食/濾過摂食のラディオドンタ類であるタミシオカリスと同じほどのサイズ(長さ0.5mm以上[9])の動物プランクトンを濾過できたが、仕組みは異なっていたと思われる。タミシオカリスの前部付属肢は密集した分岐が前後に向かって一面の濾過網を構成する[9]のに対して、エーギロカシスの前部付属肢の繊毛は全て内側に向かっており、前後で多重構造の濾過装置をなしている。これによると、エーギロカシスの前部付属肢はより高い粒径範囲の物質を濾過できる[2]。頭部の巨大な甲皮も、本体が水中に前進する度に水流を前部付属肢のところへ導く機能をもっていたと思われる[2]。第7肢節内突起の比較的丈夫な分岐は、濾過摂食の際に大きすぎる物質が奥の第1-6内突起に侵入することを防ぐ役割を果たしたと考えられる[3]。
分類 編集
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ラディオドンタ類におけるエーギロカシスの系統的位置(Moysiuk & Caron 2022 に基づく)[10] |






エーギロカシス(エーギロカシス属 Aegirocassis)は模式種(タイプ種)である Aegirocassis benmoulai のみ知られている[2]。本属はフルディア、ペイトイアなどと共にフルディア科(Hurdiidae)に分類されるラディオドンタ類であり[2][6][11][12][13][14]、そのうち更にスードアングスティドントゥスなどと共にエーギロカシス亜科(Aegirocassisinae)に分類される[3]。いくつかの系統解析によると、フルディア科の中で本属はスタンレイカリス、シンダーハンネス、ペイトイアなどより派生的である同時に、フルディア、カンブロラスターなどを含んだ系統群より基盤的だとされる[12][13][14][10]。一方、直線的な内突起を基に、エーギロカシスとシンダーハンネスを近縁とする解析結果もある[7]。
食性が本属に似たとされるタミシオカリスは別系統のタミシオカリス科(Tamisiocarididae、旧称 Cetiocaridae[9])に分類されることにより、懸濁物/濾過食性はラディオドンタ類の系統内で少なくとも2回ほど独自に進化したことが示唆される[2][6][12][14]。
発見の意義 編集
ラディオドンタ類の生息期間の延長 編集
エーギロカシスは Van Roy et al. 2015 から正式に命名される[2]が、化石標本自体は、Van Roy & Briggs 2011 によって同じ堆積累層(Fezouata Formation)で見つかった他の未命名のラディオドンタ類の化石標本と共に初めて記載された[1]。それ以前では、2009年で記載されたデボン紀のシンダーハンネスはまだラディオドンタ類として認められず(本文参照)、他にカンブリア紀より先の地質時代に当たる堆積累層から確実なラディオドンタ類の化石も発見されなかった。そのため、本属含めて Van Roy & Briggs 2011 に記載されたこれらのオルドビス紀前期(約4億8,800万 - 4億7,200万年前[1])の化石標本は、ラディオドンタ類はカンブリア紀で絶滅せず、少なくとも3,000万年後のオルドビス紀前期まで存続したことを直接的に証明した[1](後にシンダーハンネスがラディオドンタ類として認められることにより、ラディオドンタ類の生息期間がデボン紀まで延長された)[9][5][6]。
胴部付属肢の対応関係 編集
原記載の Van Roy et al. 2015 では、エーギロカシスの鰭の特徴が特に注目された。これは当時までラディオドンタ類は、胴節ごとに腹側1対の鰭のみしか発見できず、「脚のあるラディオドンタ類」として一般に知られたパラペイトイアも、後に別系統の見間違いだと判明したからである。しかしこのエーギロカシスは、この体制と異なり胴節ごとに2対の鰭をもっていた[2]。


ラディオドンタ類に類縁の古生物のうち、パンブデルリオンとケリグマケラ、いわゆる「gilled lobopodians」は鰭の下に葉足動物に似たの脚(葉足)をもち、オパビニア類もおそらく似たような脚をもつと考えられる[15]。そのため、従来の知見では、ラディオドンタ類の鰭は gilled lobopodians とオパビニア類の鰭に相同で、その脚はラディオドンタ類で退化消失し[16](もしくは単に化石に保存されず[17])、胴節ごとに1対の鰭しか残らなかったと考えられた。しかしエーギロカシスの発見は、この仮説を覆し、少なくともラディオドンタ類の共通祖先は胴節ごとに2対の鰭をもち、そのうち付け根が setal blades に連結した背側の退化的な鰭は gilled lobopodians とオパビニア類 の setal blades に覆われた鰭に相同で、腹側の鰭は gilled lobopodians の鰭ではなく、むしろ葉足に相同だと示唆される[2]。
この腹側の鰭と葉足の相同性は、構造上の類似性が低いという疑問点が存在する[18]が、背腹位置は対応しており、ラディオドンタ類の腹側の鰭と Gilled lobopodians の鰭の非相同性(および背側の鰭の相同性)も、逆の畳み方と setal blades の有無などの相違点に支持される[2]。また、ラディオドンタ類の腹側の鰭は往々にして発達した筋組織をもつため、パンブデルリオンが葉足のみに発達した筋肉をもつ(鰭は筋肉をもたない)ことも、この対応関係を支持する証拠とされる[19]。

エーギロカシスの発見も、節足動物の二叉型付属肢は、背側と腹側の付属肢構造の融合を通じて由来する説を支持する証拠の1つとなる。真節足動物(ラディオドンタ類などより派生的な節足動物)の二叉型付属肢のうち、外葉(exite)と内肢(endopod)はそれぞれラディオドンタ類の背側の setal blades と腹側の鰭(および Gilled lobopodians の setal blades と葉足)に相同だと考えられる[2]。これを踏まえて、背腹の付属肢構造(setal blades を有する背側の鰭とそれを欠く腹側の鰭)が二叉型に融合しない形質も、ライララパクスから得られる神経解剖学的証拠(少数な脳神経節)[20]に併せて、ラディオドンタ類は真節足動物に含まれるもの[21]や汎節足動物以外の脱皮動物(環神経動物)に類縁するもの[22]ではなく、基盤的な節足動物(ステムグループ節足動物)である説を支持する証拠とされる[2]。
| 分類群 | 背側の付属肢要素 | 腹側の付属肢要素 |
|---|---|---|
| 葉足動物 | - | 葉足 |
| Gilled lobopodians、オパビニア類 | Setal blades 鰭 |
葉足 |
| (従来の知見) ラディオドンタ類 |
Setal blades 鰭 |
(退化) |
| (本文の知見) ラディオドンタ類: エーギロカシスなど(フルディア科) |
Setal blades 背側の鰭 |
腹側の鰭 |
| (本文の知見) ラディオドンタ類: アノマロカリス科、アンプレクトベルア科 |
Setal blades | 鰭 |
| 真節足動物 | 外葉 | 内肢 |
ラディオドンタ類の体制への再検討 編集
-
 エーギロカシスの発見以前の知見に基づいたペイトイアの復元模型。胴節ごとに1対の鰭のみをもつとされる。
エーギロカシスの発見以前の知見に基づいたペイトイアの復元模型。胴節ごとに1対の鰭のみをもつとされる。 -
 エーギロカシスの原記載に行われる再検証により背側の鰭の存在が判明し、その発見を反映したペイトイアの復元図。
エーギロカシスの原記載に行われる再検証により背側の鰭の存在が判明し、その発見を反映したペイトイアの復元図。 -




エーギロカシスの発見により、従来のラディオドンタ類も背側の鰭があるかは再検証すべきとなった。例えばペイトイアの場合、従来の一部の化石標本から大きな鰭の基部と重なった小さな鰭の存在が確認され、これが背側鰭だと考えられる[2]。一部のフルディアの化石標本からも、背腹2対の鰭と思われる痕跡が見られる[2]。この2属は、いずれもエーギロカシスと同じくフルディア科に分類されるラディオドンタ類である(前述参照)。一方、アノマロカリス科(アノマロカリスなど)とアンプレクトベルア科(アンプレクトベルアなど)からは背側の鰭が発見できなかった。そのため、これらのラディオドンタ類の背側の鰭は単に化石に保存されず、もしくは胴部のものが二次的に退化して setal blades のみを残し、尾部のものが尾扇を構成する尾鰭に特化したと考えられる[2]。これにより、背側の鰭はラディオドンタ類の中でフルディア科に特有の派生形質である可能性も示唆される[2][10]。
また、鰭の脈と鰓らしき櫛状構造「setal blades」の構成も、本属の発見により新たな情報を得られていた。従来、鰭の脈は途中に分節しないロード状の構造で[23]、setal blades は体節の前縁に沿って繋がる部分だと考えられた[1]。しかし本属の発見により、鰭の脈は数多くの管状構造に細分され、setal blades は胴節の前縁ではなく、両背側に付いた背側の鰭の付け根のみに連結し、そこからお互いに背側の正中線に向かって伸びていた(エーギロカシスとペイトイアの場合は更に左右融合した)附属体であることが示唆される[2]。
ラディオドンタ類のオルドビス紀の生態との関係性 編集
オルドビス紀の堆積累層からエーギロカシスのような大型濾過摂食者が見つかることにより、当時における生物多様性が急速に上昇する現象「オルドビス紀生物大放散事変」(Great Ordovician Biodiversification Event, GOBE)は、プランクトンからなる生態系までにも及ぶことが示唆される[2][18]。また、同じ Fezouata Formation に産するラディオドンタ類の化石のうちフルディア科の種類のみ知られ(アノマロカリス科やアンプレクトベルア科のような獰猛な捕食者が全く見当たらない)、しかもエーギロカシスやスードアングスティドントゥスのような濾過摂食性の種類がほとんどであるため、ラディオドンタ類はフルディア科のみオルドビス紀に生き延びて、そして前述した大放散事変の影響により濾過摂食の種類が優勢になったことも示される[3]。
脚注 編集
- ^ a b c d e f g h Van Roy, Peter; Briggs, Derek E. G. (2011-05). “A giant Ordovician anomalocaridid” (英語). Nature 473 (7348): 510–513. doi:10.1038/nature09920. ISSN 0028-0836.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay az ba bb bc bd be bf bg bh bi Van Roy, Peter; Daley, Allison C.; Briggs, Derek E. G. (2015). “Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps”. Nature 522 (7554): 77–80. doi:10.1038/nature14256. ISSN 0028-0836. PMID 25762145.
- ^ a b c d e f g Potin, Gaëtan J.-M.; Gueriau, Pierre; Daley, Allison C. (2023). “Radiodont frontal appendages from the Fezouata Biota (Morocco) reveal high diversity and ecological adaptations to suspension-feeding during the Early Ordovician”. Frontiers in Ecology and Evolution 11. doi:10.3389/fevo.2023.1214109/full. ISSN 2296-701X.
- ^ a b Van Roy, Peter; Briggs, Derek E. G.; Gaines, Robert R. (2015). “The Fezouata fossils of Morocco; an extraordinary record of marine life in the Early Ordovician”. Journal of the Geological Society 172: 2015–017. doi:10.1144/jgs2015-017. ISSN 0016-7649.
- ^ a b Zeng, Han; Zhao, Fangchen; Yin, Zongjun; Zhu, Maoyan (2017). “Morphology of diverse radiodontan head sclerites from the early Cambrian Chengjiang Lagerstätte, south-west China”. Journal of Systematic Palaeontology 16 (1): 1–37. doi:10.1080/14772019.2016.1263685. ISSN 1477-2019.
- ^ a b c d e f Lerosey-Aubril, Rudy; Pates, Stephen (2018-09-14). “New suspension-feeding radiodont suggests evolution of microplanktivory in Cambrian macronekton” (英語). Nature Communications 9 (1). doi:10.1038/s41467-018-06229-7. ISSN 2041-1723.
- ^ a b Zeng, Han; Zhao, Fangchen; Zhu, Maoyan (2022-09-07). “Innovatiocaris , a complete radiodont from the early Cambrian Chengjiang Lagerstätte and its implications for the phylogeny of Radiodonta”. Journal of the Geological Society. doi:10.1144/jgs2021-164. ISSN 0016-7649.
- ^ a b c d e Pates, Stephen; Lerosey-Aubril, Rudy; Daley, Allison C.; Kier, Carlo; Bonino, Enrico; Ortega-Hernández, Javier (2021-01-19). “The diverse radiodont fauna from the Marjum Formation of Utah, USA (Cambrian: Drumian)” (英語). PeerJ 9: e10509. doi:10.7717/peerj.10509. ISSN 2167-8359.
- ^ a b c d Vinther, Jakob; Stein, Martin; Longrich, Nicholas R.; Harper, David A. T. (2014-03). “A suspension-feeding anomalocarid from the Early Cambrian” (英語). Nature 507 (7493): 496–499. doi:10.1038/nature13010. ISSN 0028-0836.
- ^ a b c Moysiuk, Joseph; Caron, Jean-Bernard (2022-07-08). “A three-eyed radiodont with fossilized neuroanatomy informs the origin of the arthropod head and segmentation” (English). Current Biology 0 (0). doi:10.1016/j.cub.2022.06.027. ISSN 0960-9822. PMID 35809569.
- ^ Liu, Jianni; Lerosey-Aubril, Rudy; Steiner, Michael; Dunlop, Jason A; Shu, Degan; Paterson, John R (2018-06-01). “Origin of raptorial feeding in juvenile euarthropods revealed by a Cambrian radiodontan”. National Science Review 5 (6): 863–869. doi:10.1093/nsr/nwy057. ISSN 2095-5138.
- ^ a b c Moysiuk, J.; Caron, J.-B. (2019-08-14). “A new hurdiid radiodont from the Burgess Shale evinces the exploitation of Cambrian infaunal food sources”. Proceedings of the Royal Society B: Biological Sciences 286 (1908): 20191079. doi:10.1098/rspb.2019.1079. PMC 6710600. PMID 31362637.
- ^ a b Moysiuk, Joseph; Caron, Jean-Bernard (2021-05-17). “Exceptional multifunctionality in the feeding apparatus of a mid-Cambrian radiodont” (英語). Paleobiology: 1–21. doi:10.1017/pab.2021.19. ISSN 0094-8373.
- ^ a b c Caron, J.-B.; Moysiuk, J. (2021-09-08). “A giant nektobenthic radiodont from the Burgess Shale and the significance of hurdiid carapace diversity”. Royal Society Open Science 8 (9): 210664. doi:10.1098/rsos.210664.
- ^ Budd, Graham E.; Daley, Allison C. (2011). “The lobes and lobopods of Opabinia regalis from the middle Cambrian Burgess Shale: The lobes of Opabinia” (英語). Lethaia 45 (1): 83–95. doi:10.1111/j.1502-3931.2011.00264.x.
- ^ C., Daley, Allison (2010). “The morphology and evolutionary significance of the anomalocaridids”. DIVA.
- ^ Xianguang, Hou; Jan, Jan Bergström; Jiayu, In Rong; Zongjie, Fang; Zhanghe, Zhou; Renbin, Zhan; Xiangdong, Wang; (eds, Yuan Xunlai et al. (2006). Dinocaridids – anomalous arthropods or arthropod-like worms?.
- ^ a b “Palaeontology: In a Flap About Flaps” (英語). Current Biology 25 (12): R503–R506. (2015-06-15). doi:10.1016/j.cub.2015.04.029. ISSN 0960-9822.
- ^ Young, Fletcher J.; Vinther, Jakob (2016-11-25). “Onychophoran-like myoanatomy of the Cambrian gilled lobopodian Pambdelurion whittingtoni” (英語). Palaeontology 60 (1): 27–54. doi:10.1111/pala.12269. ISSN 0031-0239.
- ^ Cong, Peiyun; Ma, Xiaoya; Hou, Xianguang; Edgecombe, Gregory D.; Strausfeld, Nicholas J. (2014-07-16). “Brain structure resolves the segmental affinity of anomalocaridid appendages” (英語). Nature 513 (7519): 538–542. doi:10.1038/nature13486. ISSN 0028-0836.
- ^ Haug, Joachim; Waloszek, Dieter; Maas, Andreas; Liu, Yu; Haug, Carolin (2012-03-01). “Functional morphology, ontogeny and evolution of mantis shrimp-like predators in the Cambrian”. Palaeontology 55: 369–399. doi:10.1111/j.1475-4983.2011.01124.x.
- ^ Xian‐Guang, Hou; Bergström, Jan; Ahlberg, Per (1995-09-01). “Anomalocaris and Other Large Animals in the Lower Cambrian Chengjiang Fauna of Southwest China”. GFF 117: 163–183. doi:10.1080/11035899509546213.
- ^ Daley, Allison; R Paterson, John; Edgecombe, Gregory; García-Bellido, Diego; Jago, James (2013-03-01). “New anatomical information on Anomalocaris from the Cambrian Emu Bay Shale of South Australia and a reassessment of its inferred predatory habits”. Palaeontology 56: 971–990. doi:10.1111/pala.12029.
関連項目 編集
外部リンク 編集
- Aegirocassis benmoulae - YouTube - イェール・ピーボディ自然史博物館によるエーギロカシスの紹介映像(英語)