麻痺性貝毒を産生する渦鞭毛藻。日本近海では G. catenatum Graham などが出現する。従前は Karenia 属がここに含まれていた。
Karenia 属
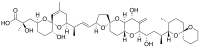
神経性貝毒(neurotoxic shellfish poisoning; NSP)を産生する渦鞭毛藻。カレニア・ブレビス(K. brevis (Davis) Hansen et Moestrup)や K. mikimotoi (Mikyake et Kominami ex Oda) Hansen et Moestrup が代表的である。毒はブレベトキシンである。本属は以前は Gymonodinium 属に含められていたが、2000年に分離された[8]。
K. brevis による貝毒の被害は北米とニュージーランドで発生している。前者では魚およびマナティーの大量死が報告されている。後者では1992年に300名近い食中毒患者を出した[9]。日本では東京湾などで K. brevis、K. mikimotoi ともに確認されている[10][11]ものの、食中毒の発生報告はいまだない。
また、カレニア・セリフォルミス (Karenia selliformis) は、2021年9月から11月にかけて北海道東部太平洋岸にて発生した赤潮の原因に特定されている。この赤潮によって、ウニやサケ・マス類などの斃死による大きな漁業被害が発生した。K. selliformis を原因とする赤潮が日本近海にて発生した例は、これが初めての報告となる。また、原因となった K. selliformis は、2020年のロシア・カムチャツカ半島沿岸産のものと、DNA配列が一致している。
Ostreopsis 属
O. lenticularis や O. siamensis がパリトキシンを産生する。細胞の姿形が前述の Gambierdiscus toxicus に似ているのでしばしば混同されるが、遊泳法や鎧板配列が異なることで区別できる。
Pfiesteria 属
アメリカのノースカロライナ州で発見された渦鞭毛藻。タイプ種である P. piscicida が産生する毒素はエアロゾルとして気相に放出され、人体にも影響を及ぼすと言われている。当初は Pfiesteria の正体が掴めず、生息域の近隣住民からは“phantom dinoflagellate”と呼ばれ恐れられた。P. piscicida は名前の通り魚を殺してその血球などを捕食する。毒素は2007年に同定された[12]が名前はまだ付いていない。
同属の P. shumwayae も毒性が疑われたが、これについては毒ではなく、細胞が魚へ直接攻撃を仕掛けているのだという説もある[13][14]。いずれの Pfiesteria についても、その有毒性や生活環を巡っては様々な意見・議論があり、収束していない。詳細はフィエステリアを参照。
Prorocentrum 属
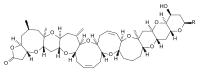
付着性の P. lima (Ehrenberg) Dodge がオカダ酸を産生する。この藻類はシガテラの発生域に多く分布しており、Gambierdiscus とともに魚の毒化との関連性が示唆されている。
Protoceratium 属
下痢性貝毒であるイェッソトキシンを産生する渦鞭毛藻。この毒素は従来 Dinophysis に由来すると思われていたが、1997年に P. reticulatum (Claparède et Lachmann) Bütschli が産生元であることが報告された。[15]
熱帯地域を中心として、麻痺性貝毒を産生する渦鞭毛藻。特に問題となる種は P. bahamense var. compressum (Böhm) Steindinger, Tester et Taylor である。発達した鎧板と明瞭な縫合線を持つ。本種はシストを形成し、海底に堆積することも報告されている[17]。
^Hallegraeff GM, Steffensen DA, Wetherbee R (1988). “Three estuarine Australian dinoflagellates theat can
produce paralytic shellfish toxins”. J. Plankton. Res.10: 533-41.
^『東京湾で観察された、渦鞭毛藻 Alexandrium minutum Halim』 水尾寛己・鳥海三郎 横浜環境科学研究所報第30号2005年 PDF available
^Matsuda A, Nishijima T, Fukami K, Adachi M (2001). “Effects of B group vitamins on the growth of toxic dinoflagellate Alexandrium catenella”. Nippon Suisan Gakkaishi67 (4): 658-63.PDF available
^Daugbjerg N, Hansen G, Larsen J, Moestrup Ø (2000). “Phylogeny of some of the major genera of dinoflagellates based on ultrastructure and partial LSU rDNA sequence data, including the erection of three new genera of unarmored dinoflagellates”. Phycologia39: 302-17.
^Bates M, Baker M, Willson N, Lane N, Handford A. (1993) Marine toxins and New Zealand shellfish. p35-40. In: Proceedings of a workshop on research issue. Royal Society of New Zealand.
^Moeller PDR, Beauchesne KR, Huncik KM, Davis WC, Christopher SJ, Riggs-Gelasco P, Gelasco AK (2007). “Metal Complexes and Free Radical Toxins Produced by Pfiesteria piscicida”. Environ. Sci. Technol.41 (4): 1166-72.doi:10.1021/es0617993
^Berry JP, Reece KS, Rein KS, Baden DG, Haas LW, Ribeiro WL, Shields JD, Snyder RV, Vogelbein KW, Gawley RE (2002). “Are Pfiesteria species toxicogenic? Evidence against production of ichthyotoxins by Pfiesteria shumwayae”. PNAS99 (17): 10970-5.doi:10.1073/pnas.172221699
^Vogelbein WK, Lovko VJ, Shields JD, Reece KS, Mason PL, Haas LW, Walker CC (2002). “Pfiesteria shumwayae kills fish by micropredation not exotoxin secretion”. Nature418: 967-970.doi:10.1038/nature01008
^Satake M, MacKenzie L, Yasumoto T (1997). “Identification of Protoceratium reticulatum as the biogenetic origin of yessotoxin”. Natural toxins5: 164-7.
^James KJ, Moroney C, Roden C, Satake M, Yasumoto T, Lehane M, Furey A (2003). “Ubiquitous 'benign' alga emerges as the cause of shellfish contamination responsible for the human toxic syndrome, azaspiracid poisoning”. Toxicon41 (2): 145-51.PMID12565733
^Matsuoka K, Fukuyo Y, Praseno FD, Adnan Q, Kodama M. (1999) Dinoflagellate Cysts in Surface Sediments of Jakarta Bay, off Ujung Pandang and Larantuka of Flores Islands, Indonesia with Special Reference of Pyrodinium bahamense. 長崎大学水産学部 研究報告 80号 他
^Lin Y-Y, Risk M, Ray SM, Van Engen D, Clardy J, Golik J, James JC, Nakanishi K (1981). “Isolation and structure of brevetoxin B from the “Red Tide” dinoflagellate ptychodiscus brevis (Gymnodinium breve)”. J. Am. Chem. Soc.103: 6773–5.doi:10.1021/ja00412a053
^Ito K, Toyoda I, Higashiyama M, Uemura D, Sato MH, Yoshimura SH, Ishii T, Takeyasu K (2003). “Channel induction by palytoxin in yeast cells expressing Na+,K+-ATPase or its chimera with sarco/endoplasmic reticulum Ca2+-ATPase”. FEBS Lett.543: 108-12.
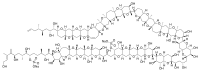
^Sasaki M, Matsumori N, Maruyama T, Nonomura T, Murata M, Tachibana K, Yasumoto T (1996). “The complete structure of maitotoxin, part I: configuration of the C1—C14 side chain.”. Angew. Chem. Int. Ed. Engl.35: 1672–5.doi:10.1002/anie.199616721
^Nonomura T, Sasaki M, Matsumori N, Murata M, Tachibana K, Yasumoto T (1996). “The complete structure of maitotoxin, part II: configuration of the C135—C142 side chain and absolute configuration of the entire molecule”. Angew. Chem. Int. Ed. Engl.35: 1675–8.doi:10.1002/anie.199616751
^『赤潮対策(有害プランクトンの早期識別)について』 長崎県総合水産試験場 漁連だより133号 2006年5月 PDF available
^Sako Y, Hosoi-Tanabe S, Uchida A (2004). “Fluorescence in situ Hybridization Using rRNA-Targeted Probes for Simple and Rapid Identification of the Toxic Dinoflagellates Alexandrium tamarense and Alexandrium catenella”. Journal of Phycology40 (3): 598-605.